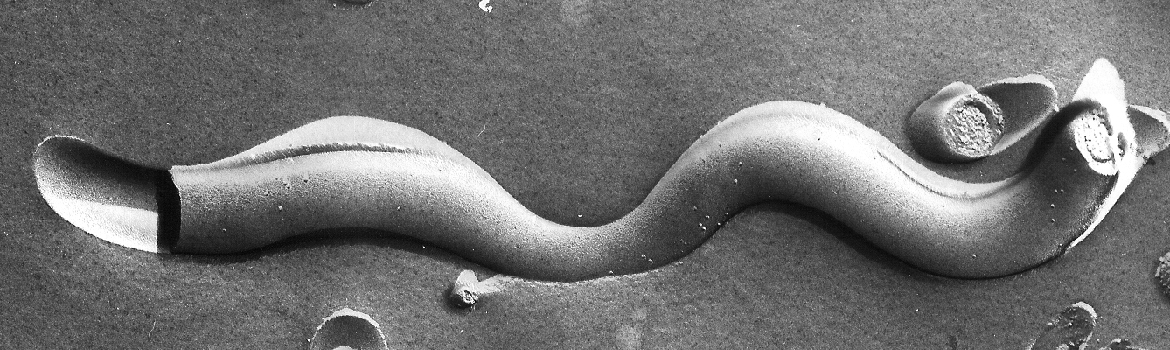
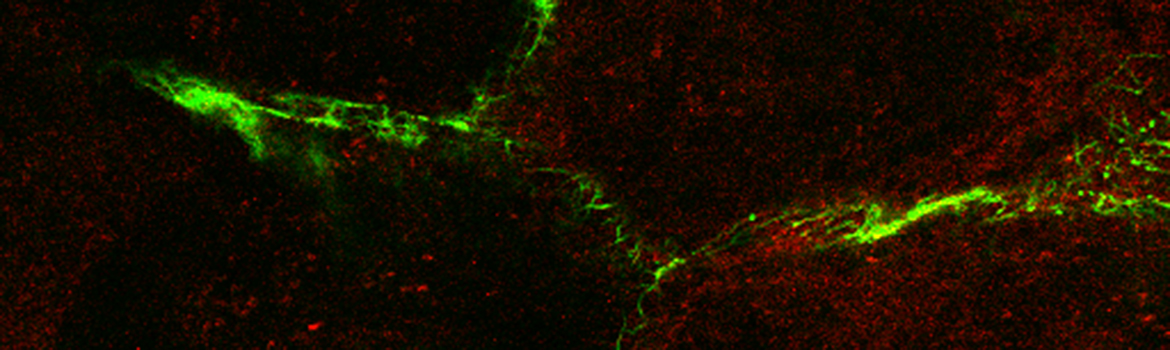
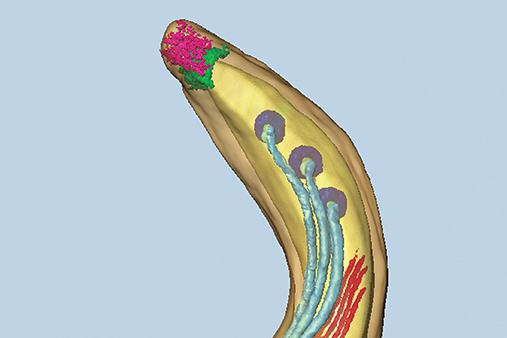
Spirochetes are an ancient and extremely successful eubacterial phylum characterized by distinctive helical or planar wave-form morphology and flagellar filaments confined to the periplasmic space. Spirochetes from the genera Leptospira, Treponema, and Borrelia are highly invasive pathogens that, as the agents of leptospirosis, syphilis (T. pallidum), Lyme disease (B. burgdorferi), and relapsing fever (B. hermsii, B. recurrentis, and others), pose public health problems of global dimensions. T. denticola and numerous other treponemal species, most of which remain uncultivated, are major components of the polymicrobial biofilms that cause periodontal disease, the major cause of tooth loss worldwide. In recent years, the availability of genomic sequences, improved methodologies for genetically manipulating these fastidious bacteria, and novel light and microscopic technologies have provided powerful new tools for studying the ultrastructures, genetics and parasitic strategies of these unusual pathogens.